หุ้น:



ส้อมการจำลองแบบคืออะไร?
ส้อมการจำลองแบบ มันเป็นจุดที่มีการจำลองดีเอ็นเอเกิดขึ้นมันถูกเรียกว่าจุดเติบโต มันมีรูปร่าง Y และเมื่อมีการจำลองแบบกิ๊บจะถูกแทนที่ด้วยโมเลกุลของดีเอ็นเอ.
การจำลองดีเอ็นเอเป็นกระบวนการของเซลล์ที่เกี่ยวข้องกับการทำสำเนาของสารพันธุกรรมในเซลล์ โครงสร้างของ DNA เป็นเกลียวคู่และในการทำซ้ำเนื้อหาจะต้องเปิด แต่ละเส้นจะเป็นส่วนหนึ่งของห่วงโซ่ DNA ใหม่เนื่องจากการจำลองแบบเป็นกระบวนการกึ่งอคติ.
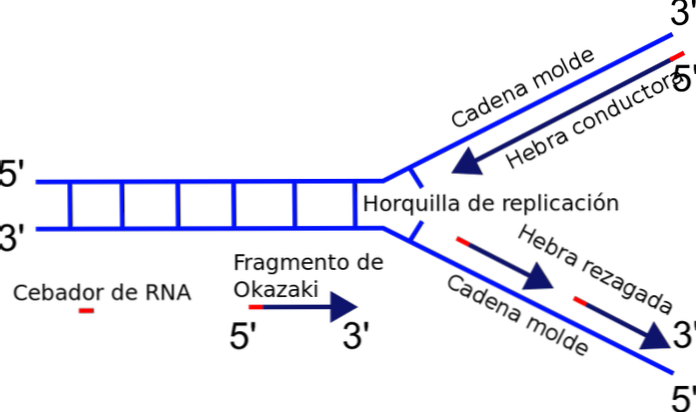
ส้อมเรพลิเคชันจะเกิดขึ้นระหว่างทางแยกระหว่างเทมเพลตหรือแม่แบบที่แยกจากกันใหม่และเทมเพลต DNA ที่ยังไม่ได้ทำซ้ำ เมื่อเริ่มต้นการจำลองดีเอ็นเอเส้นหนึ่งสามารถทำซ้ำได้อย่างง่ายดายในขณะที่คนอื่น ๆ ต้องเผชิญกับปัญหาขั้ว.
เอ็นไซม์ที่รับผิดชอบในการทำโพลิเมอไรเซชั่นในห่วงโซ่ - DNA polymerase - สังเคราะห์เฉพาะดีเอ็นเอของเกลียวในทิศทาง 5'-3 ' ดังนั้นเส้นหนึ่งต่อเนื่องและอีกสายหนึ่งทนต่อการจำลองแบบไม่ต่อเนื่องสร้างชิ้นส่วนของ Okazaki.
ดัชนี
- 1 การจำลองแบบของดีเอ็นเอและส้อมการจำลองแบบ
- 1.1 การจำลองแบบทิศทางเดียวและสองทิศทาง
- 1.2 เอนไซม์ที่เกี่ยวข้อง
- 1.3 จุดเริ่มต้นของการจำลองแบบและการสร้างทางแยก
- 1.4 การยืดตัวและการเคลื่อนที่ของทางแยก
- 1.5 การสิ้นสุด
- 2 การจำลองแบบดีเอ็นเอเป็นแบบอสังหา
- 3 ปัญหาขั้วไฟฟ้า
- 3.1 วิธีการทำงานของโพลีเมอเรส?
- 3.2 การผลิตชิ้นส่วนโอกาซากิ
- 4 อ้างอิง
การจำลองแบบของดีเอ็นเอและส้อมการจำลองแบบ
DNA เป็นโมเลกุลที่เก็บข้อมูลทางพันธุกรรมที่จำเป็นสำหรับสิ่งมีชีวิตทุกชนิดยกเว้นไวรัสบางชนิด.
พอลิเมอร์ขนาดใหญ่นี้ประกอบด้วยนิวคลีโอไทด์ที่แตกต่างกันสี่ (A, T, G และ C) อยู่ในนิวเคลียสของยูคาริโอตในแต่ละเซลล์ที่ประกอบขึ้นเป็นเนื้อเยื่อของสิ่งมีชีวิตเหล่านี้ (ยกเว้นในเซลล์เม็ดเลือดแดงของสัตว์เลี้ยงลูกด้วยนม ของแกน).
ทุกครั้งที่เซลล์แบ่งดีเอ็นเอจะต้องทำซ้ำเพื่อให้กำเนิดเซลล์ลูกสาวด้วยวัสดุทางพันธุกรรม.
การจำลองแบบทิศทางเดียวและแบบสองทิศทาง
การจำลองแบบสามารถเป็นทิศทางเดียวหรือแบบสองทิศทางขึ้นอยู่กับการสร้างส้อมการจำลองที่จุดกำเนิด.
ในกรณีของการจำลองแบบในทิศทางเดียวจะเกิดส้อมเดียวเท่านั้นในขณะที่การจำลองแบบสองทางสองส้อมจะเกิดขึ้น.
เอนไซม์ที่เกี่ยวข้อง
สำหรับกระบวนการนี้จำเป็นต้องใช้เครื่องจักรที่ซับซ้อนของเอนไซม์ซึ่งทำงานได้อย่างรวดเร็วและสามารถจำลองดีเอ็นเอได้อย่างแม่นยำ เอนไซม์ที่สำคัญที่สุดคือ DNA polymerase, DNA primase, DNA helicase, DNA ligase และ topoisomerase.
จุดเริ่มต้นของการจำลองแบบและการก่อตัวของส้อม
การจำลองดีเอ็นเอไม่ได้เริ่มต้นที่สถานที่สุ่มใด ๆ ในโมเลกุล มีภูมิภาคเฉพาะใน DNA ที่ทำเครื่องหมายจุดเริ่มต้นของการจำลองแบบ.
ในแบคทีเรียส่วนใหญ่โครโมโซมแบคทีเรียมีจุดเริ่มต้นที่อุดมไปด้วย AT เดียว องค์ประกอบนี้มีเหตุผลเนื่องจากช่วยอำนวยความสะดวกในการเปิดพื้นที่ (คู่ AT จะเข้าร่วมโดยสะพานไฮโดรเจนสองแห่งในขณะที่คู่ GC สามคน).
เมื่อ DNA เริ่มเปิดขึ้นโครงสร้างของรูปตัว Y จะเป็นตัวแยกการจำลองแบบ.
การยืดตัวและการเคลื่อนไหวของทางแยก
DNA polymerase ไม่สามารถเริ่มการสังเคราะห์โซ่ลูกตั้งแต่เริ่มต้นได้ คุณต้องมีโมเลกุลที่มี 3'-end เพื่อให้โพลีเมอเรสมีจุดเริ่มต้นในการเกิดพอลิเมอร์.
ปลายฟรี 3 'นี้เสนอโดยนิวคลีโอไทด์โมเลกุลขนาดเล็กที่เรียกว่าไพรเมอร์หรือไพรเมอร์ ครั้งแรกที่ทำหน้าที่เป็นตะขอสำหรับโพลิเมอร์.
ด้วยหลักสูตรการจำลองแบบส้อมการจำลองมีความสามารถในการเคลื่อนที่ไปตาม DNA ระยะห่างของส้อมการทำซ้ำใบโมเลกุล DNA สองวงสองวงที่กำหนดการก่อตัวของโมเลกุลลูกสาวสองวง.
ทางแยกสามารถเพิ่มขึ้นได้ด้วยการกระทำของเอนไซม์เฮลิเคสที่คลายโมเลกุลโมเลกุลดีเอ็นเอ เอนไซม์นี้ทำลายพันธะไฮโดรเจนระหว่างคู่ฐานและช่วยให้การกระจัดของส้อม.
เสร็จสิ้น
การจำลองแบบถูกยกเลิกเมื่อส้อมทั้งสองอยู่ที่ 180 ° C จากจุดกำเนิด.
ในกรณีนี้เราพูดถึงว่ากระบวนการจำลองแบบในแบคทีเรียไหลและจำเป็นต้องเน้นกระบวนการบิดทั้งหมดของโมเลกุลวงกลมที่เกี่ยวข้องกับการจำลองแบบ Topoisomerase มีบทบาทสำคัญในการคลายโมเลกุล.
การจำลองดีเอ็นเอเป็นแบบอสังหา
คุณเคยสงสัยหรือไม่ว่าการจำลองแบบเกิดขึ้นใน DNA ได้อย่างไร? กล่าวคือเกลียวคู่อีกอันต้องเกิดขึ้นจากเกลียวคู่ แต่มันเกิดขึ้นได้อย่างไร เป็นเวลาหลายปีนี่เป็นคำถามเปิดในหมู่นักชีววิทยา อาจมีหลายวิธีเรียงสับเปลี่ยน: สองเส้นเก่าด้วยกันและสองใหม่เข้าด้วยกันหรือด้ายใหม่และเก่าเพื่อสร้างเกลียวคู่.
ในปี 1957 นักวิจัย Matthew ตอบคำถามนี้และ Franklin Stahl รูปแบบการจำลองแบบที่เสนอโดยผู้เขียนคือเซมิคอนเซอร์โวทีฟ.
Meselson และ Stahl ระบุว่าผลของการจำลองแบบเป็นโมเลกุลดีเอ็นเอสองเส้นที่ควั่น โมเลกุลที่เกิดขึ้นแต่ละอันประกอบด้วยเส้นเก่า (จากแม่หรือโมเลกุลเริ่มต้น) และสายใหม่ที่สังเคราะห์ขึ้นใหม่.
ปัญหาขั้วไฟฟ้า
วิธีการทำงานของโพลีเมอเรส?
DNA helix ประกอบด้วยสองโซ่ที่วิ่งไปในทิศทางตรงกันข้าม: หนึ่งไปในทิศทาง 5'-3 'และอีก 3'-5'.
เอนไซม์ที่สำคัญที่สุดในกระบวนการจำลองคือ DNA polymerase ซึ่งมีหน้าที่ในการเร่งการจับตัวของนิวคลีโอไทด์ใหม่ที่จะถูกเพิ่มเข้าไปในห่วงโซ่ DNA polymerase สามารถขยายสายโซ่ในทิศทาง 5'-3 'เท่านั้น ความจริงเรื่องนี้เป็นอุปสรรคต่อการทำซ้ำของโซ่ในการแยกการจำลองแบบพร้อมกัน.
ทำไม? การเพิ่มขึ้นของนิวคลีโอไทด์เกิดขึ้นที่ฟรีเอนด์ 3'where พบกลุ่มไฮดรอกซิล (-OH) ดังนั้นมีเพียงโซ่เดียวเท่านั้นที่สามารถขยายได้อย่างง่ายดายโดยการเติมขั้วของนิวคลีโอไทด์จนถึงปลาย 3 ' สิ่งนี้เรียกว่าเส้นใยนำไฟฟ้าหรือต่อเนื่อง.
การผลิตชิ้นส่วนโอกาซากิ
สายอื่น ๆ ไม่สามารถยืดตัวได้เนื่องจากปลายอิสระคือ 5 'และไม่ใช่ 3' และไม่มีโพลิเมอร์ทำให้เร่งการเพิ่มของนิวคลีโอไทด์จนถึงปลาย 5 ' ปัญหาได้รับการแก้ไขโดยการสังเคราะห์ชิ้นส่วนสั้นหลายชิ้น (130 ถึง 200 นิวคลีโอไทด์) แต่ละชิ้นในทิศทางปกติของการจำลองจาก 5 'ถึง 3'.
การสังเคราะห์ชิ้นส่วนที่ไม่ต่อเนื่องนี้สิ้นสุดลงด้วยการรวมกันของแต่ละส่วนปฏิกิริยาที่เร่งปฏิกิริยาโดย DNA ligase เพื่อเป็นเกียรติแก่ผู้ค้นพบกลไกนี้ Reiji Okazaki กลุ่มเล็ก ๆ ที่ถูกสังเคราะห์เรียกว่าชิ้นส่วนของ Okazaki.
การอ้างอิง
- Alberts, B. , Bray, D. , Hopkin, K. , Johnson, A.D. , Lewis, J. , Raff, M. , ... & Walter, P. (2015). ชีววิทยาของเซลล์ที่สำคัญ. วิทยาศาสตร์พวงมาลัย.
- Cann, I. K. , & Ishino, Y. (1999) การจำลองดีเอ็นเอ Archaeal: ระบุชิ้นส่วนเพื่อแก้ปริศนา. พันธุศาสตร์, 152(4), 1249-67.
- Cooper, G. M. , & Hausman, R. E. (2004). เซลล์: วิธีโมเลกุล. Medicinska naklada.
- Garcia-Diaz, M. , & Bebenek, K. (2007) ฟังก์ชั่นหลากหลายของ DNA polymerases. บทวิจารณ์ที่สำคัญในสาขาวิทยาศาสตร์พืช, 26(2), 105-122.
- Lewin, B. (2008). ทรงเครื่อง. Mc Graw-Hill Interamericana.
- Shcherbakova, P. V. , Bebenek, K. , & Kunkel, T. A. (2003) หน้าที่ของโพลิเมอร์ยูคาริโอต DNA. วิทยาศาสตร์ของ SAGE KE, 2003(8), 3.
- Steitz, T. A. (1999) DNA polymerases: ความหลากหลายของโครงสร้างและกลไกทั่วไป. วารสารเคมีชีวภาพ, 274(25), 17395-17398.
- วัตสัน, J. D. (2006). อณูชีววิทยาของยีน. Ed. Panamericana การแพทย์.
- Wu, S. , เครา, W. A. , Pedersen, L. G. , & Wilson, S. H. (2013) การเปรียบเทียบโครงสร้างของสถาปัตยกรรมดีเอ็นเอพอลิเมอร์แสดงให้เห็นว่าเกตเวย์นิวคลีโอไทด์ไปยังไซต์แอคทีฟโพลิเมอร์. รีวิวเคมี, 114(5), 2759-74.